提起miRNA与mRNA互作,大家首先想到的结合区域是不是mRNA的3′UTR区?并且还能朗朗上口地告诉小医,该实验的关键就是将待测mRNA的3′UTR序列插入报告基因载体,再共转入对应的miRNA,如果荧光素酶活性下降,则提示为其靶序列。
那么,你知道吗?其实在mRNA的蛋白编码区(CDS区)也存在miRNA结合位点,并且近年来,越来越多的证据证实许多功能性miRNA靶位点位于哺乳动物mRNA转录本的CDS区,而当靶点位于哺乳动物的CDS区时,位点可及性和局部翻译效率对于miRNA的作用十分重要(图1)。
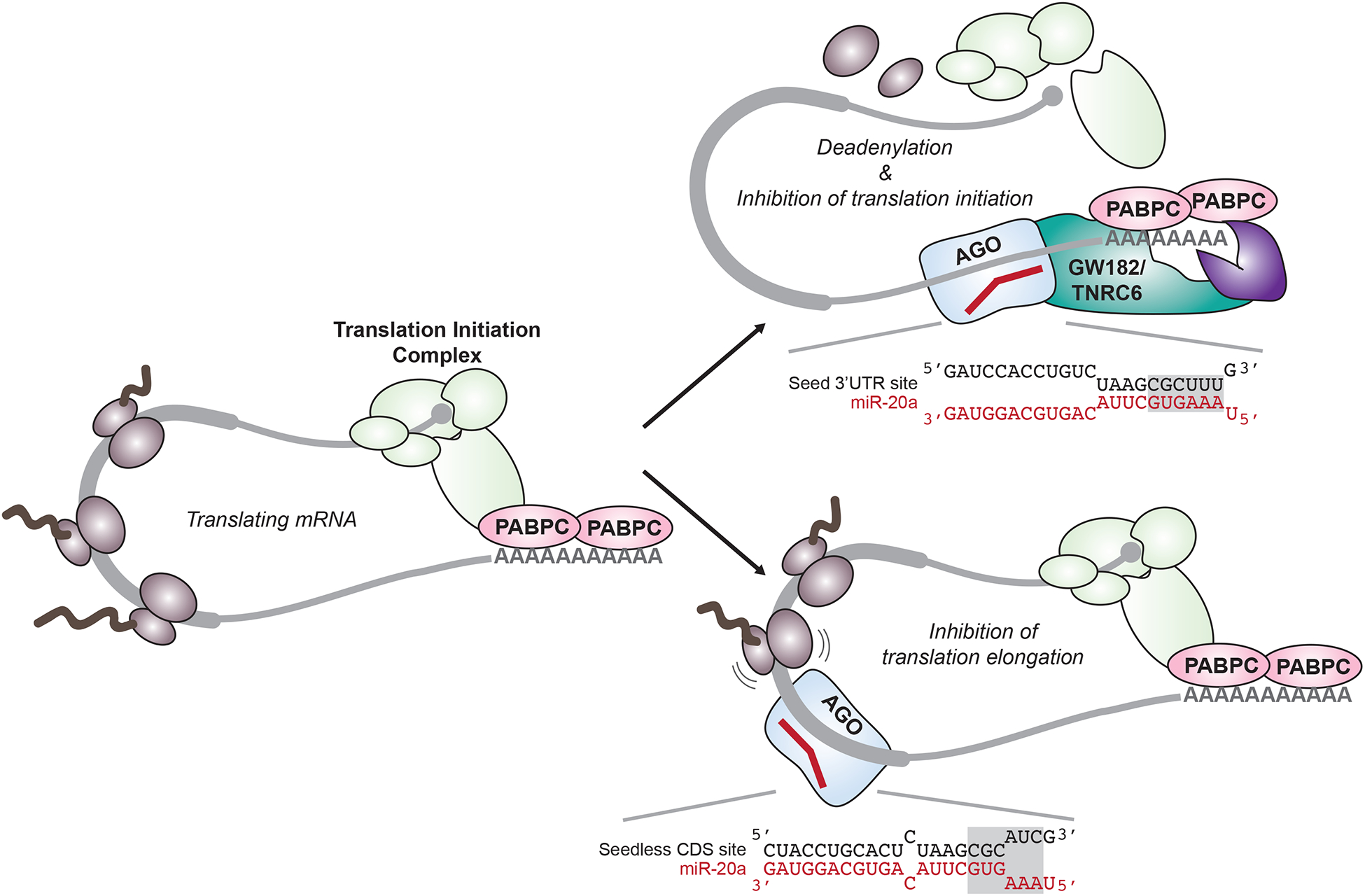
接下来,就和小医一起来了解下miRNA与mRNA靶位点的研究历史吧!
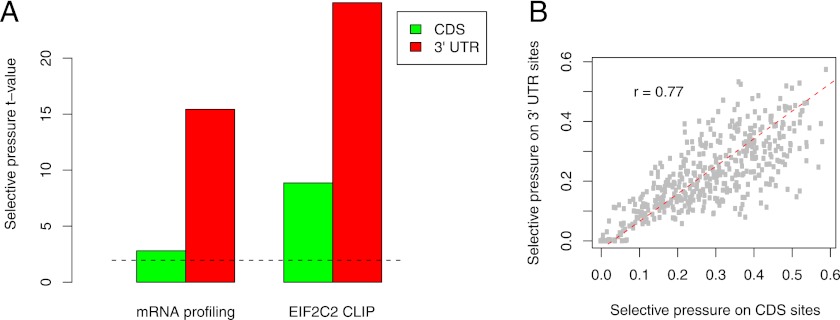
基于进化保守的方法预测CDS区中的miRNA靶位点,结果显示,预测CDS位点最多的miRNA包括hsa-let-7a-5p、hsa-miR-9-5p、hsa-miR-125a-5p和hsa-miR-153(Hausser et al., 2013)。介于ElMMo是可用的最准确的miRNA靶点预测方法之一,Hausser等人利用ElMMo来预测CDS区中的miRNA靶位点,分析了八个已发布的数据集,并比较了单个miRNA转染前后的mRNA表达量,结果表明ElMMo不仅可以预测3’UTR中的功能位点,还可以预测CDS中的功能位点,且CDS和3’UTR位点是平行进化的,但CDS位点具有在进化中赋予选择优势的功能(图2)。

期刊:Nature Structural & Molecular Biology
发表时间:2018.10.8
影响因子:16.8
技术手段:RT-qPCR,荧光素酶报告基因检测实验,核糖体分析(Ribo-seq),RNA-seq,ChIP-qPCR,eCLIP-seq等技术
DOI号:10.1038/s41594-018-0136-3
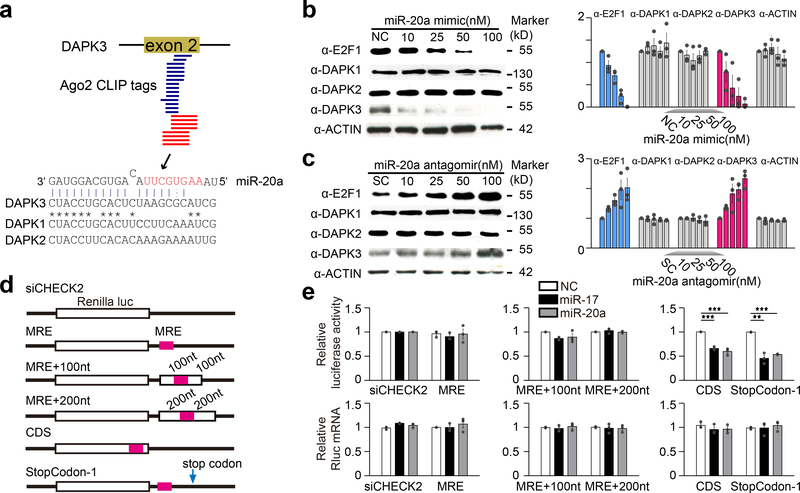
首先,作者结合其前期Ago2 CLIP-seq数据及文献调研,通过CLASH作图确定HEK293T细胞中与miR-20a互作的DAPK3位点(图4a)。进一步的Western Blot实验结果表明,DAPK3蛋白水平被miR-20a mimic特异性抑制,并被miR-20a antagomir升高(图4b,c)。
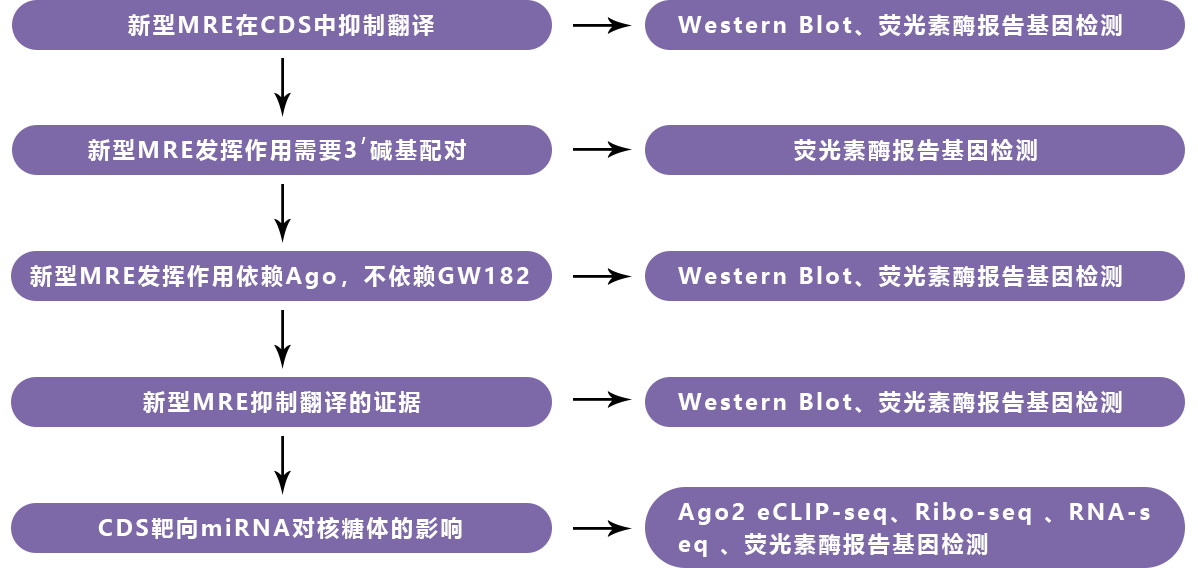
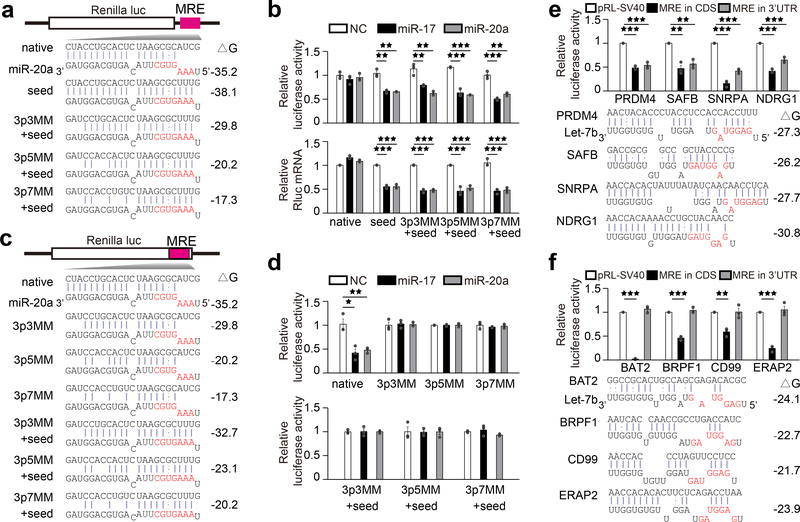
2、CDS MRE需要广泛的3’碱基配对才能发挥作用
进一步的,作者发现,当在萤光素酶报告基因载体中引入MRE突变,重新建立种子碱基配对时(图5a),3’UTR中的MRE发挥作用(图5b)。为了确定3’碱基配对对于MRE在CDS中的作用是否至关重要,在MRE中引入了突变,发现3’碱基配对确实对于MRE发挥作用至关重要,即使在恢复种子碱基配对后也是如此(图5c、d)。
接下来,作者通过挖掘CLASH数据,确定了两类假定MRE。第一类(即PRDM4、SAFB、SNRPA和NDRG1)显示出与Let-7b的5’和3’碱基配对潜力,并且当插入荧光素酶报告基因时,无论在3’UTR或CDS中,所有荧光素酶活性均受到抑制(图5e);第二类(即BAT2、BRPF1、CD99和ERAP2)与miR-20a MRE表现出明显的相似性,具有与Let-7b的大量3’配对潜力,但在5’处缺乏典型的碱基配对种子,当放入荧光素酶报告基因时,所有这些都只在CDS中发挥作用(图5f)。
图5 CDS和3’UTR靶向miRNA需要在3’端和5’端的种子处进行不同的碱基配对。
3、CDS靶向miRNA的功能依赖于Ago,而非GW182
由于Ago蛋白和GW182都是3’UTR区的标准MRE功能的必需因子。作者为了测试CDS区的MRE发挥功能时是否也需要这两种蛋白,分析了Ago2缺失的小鼠胚胎成纤维细胞(MEF)以及经Ago1-4 siRNA库处理的HeLa细胞中的多个非典型MRE。结果发现,这些非典型MRE的功能在Ago2缺失的MEF中部分受损,在Ago1-4敲低的HeLa细胞中完全消失,表明CDS MRE需要Ago蛋白才能发挥作用(图6a、b)。同时,作者还检测了敲低TNRC6家族成员后,CDS MRE活性及DAPK3的表达,结果并无显著差异(图6c、d)。在CDS中,DAPK3中的miR-20a MRE在3’端具有强碱基配对相互作用,但5’种子中的最小碱基配对仍然允许一定程度的Ago2招募,但不能招募GW182。
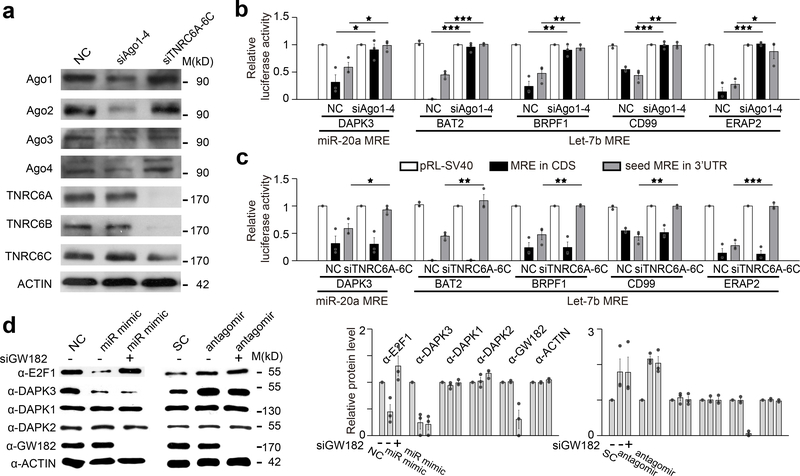
图6 CDS靶向miRNA的方式是Ago依赖性,而非GW182。
4、CDS靶向miRNA诱导翻译终止
为了研究miRNA在CDS中发挥作用时如何抑制翻译,作者利用报告基因系统探究了Let-7b对帽依赖性报告基因的抑制作用(图7a)。进一步的,作者将DAPK3的MRE插入到荧光素酶报告基因中,并将全长34kD FLAG荧光素酶蛋白截短至约28kD,为了进行比较,作者还设计了终止密码子或非功能性MRE,即在同一位置的3’端有3个碱基不匹配(图7b,上)。结果显示,野生型荧光素酶产生了预期的34kD条带,并在CDS中检测到了含有功能性MRE的报告基因的28kD条带,这种截短的产物取决于功能性MRE,在同一位置插入突变型MRE以及当细胞与miR-20a antagomir共转染时,条带显著减弱(图7b,下)。
此外,作者还采取了“2A”策略,即在GFP和MCherry之间插入P2A自切割肽序列,发现与突变型MRE相比,MCherry对来自DAPK3的野生型miR-20a MRE的反应有所减少,miR-20a过表达后进一步减少,而miR-20a antagomir减弱了这种MRE依赖性效应(图7c)。
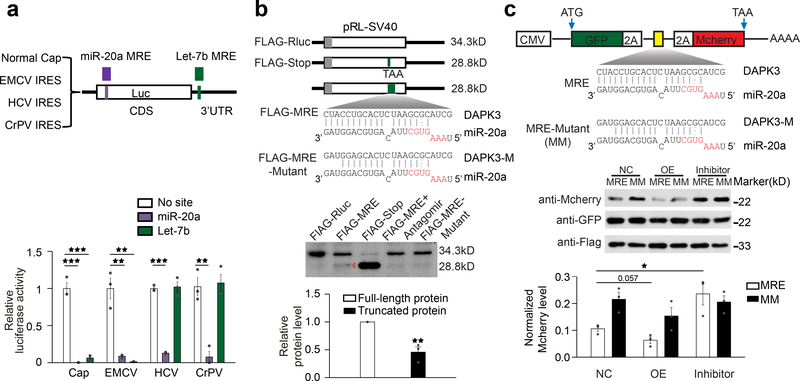
图7 CDS靶向miRNA导致翻译终止。
5、CDS靶向miRNA对核糖体结合的影响
随后,作者进行了核糖体分析,结果在miR-20a转染前后确定了特定的Ago2峰(图8a)。相对于未改变的Ago2峰或mRNA中的随机位置,CDS和3’UTR中诱导的Ago2峰与miR-20a的碱基配对潜力升高(图8b)。接下来,作者通过单个转录本的核糖体分析检验了特征化DAPK3转录本的数据,发现了Ago2的明显诱导以及特征化DAPK3转录本上RFP的适度增加,miR-20a MRE响应miR-20a过度表达,而RNA-seq谱基本保持不变(图8c)。然后,通过将RPF与miR-20a诱导的Ago2峰的中心对齐,发现,这些积累的RPF不受miR-20a过表达的影响(图8d)。此外,作者还发现含有miR-20a转录本的RNA衰变会诱导3’UTR中的Ago2峰,CDS中Ago2峰的转录本略有减少,但CDS中具有或不具有Ago2结合的转录本之间的翻译效率几乎没有差异(图8e)。
最后,作者在荧光素酶报告基因3’端附近设计了miR-20a MRE,但仍在CDS区域内,发现在同一位置放置终止密码子不会影响荧光素酶活性,表明最后一个C端序列对于荧光素酶活性来说不是必需的(图8f,左下)。相比之下,miR-20a MRE的存在降低了内源性miR-20a的荧光素酶活性,而这种效应可以用蛋白酶体抑制剂MG132来“挽救”(图8f,右下)。
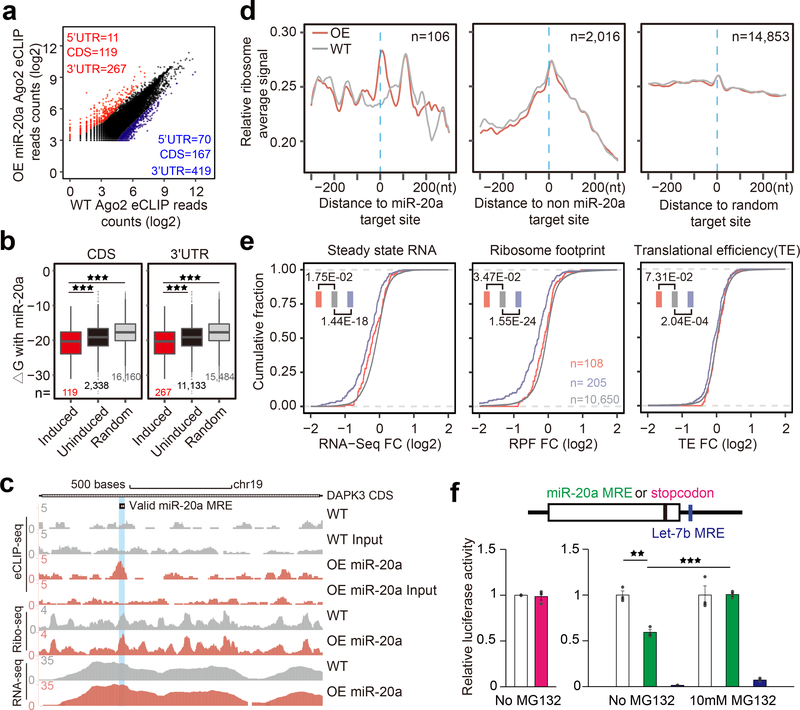
总而言之,作者通过追踪最初在p53激活激酶DAPK3的CDS区域中发现的MRE,发现了一类仅在人类CDS区域发挥作用的MRE,通过对这些“不寻常”MRE的功能和机制表征,证明CDS靶向miRNA需要在3’端而不是5’种子(miRNA 5’端2-7nt碱基为miRNA种子序列)进行的碱基配对对于miRNA识别靶标 mRNA尤为重要)中进行广泛的碱基配对;以Argonaute依赖但GW182独立的方式引起基因沉默;并通过诱导短暂的核糖体停滞而不是mRNA不稳定来抑制翻译。
Chi SW, Zang JB, Mele A, Darnell RB. Argonaute HITS-CLIP decodes microRNA-mRNA interaction maps. Nature. 2009;460(7254):479-486.
Gu W, Wang X, Zhai C, Zhou T, Xie X. Biological basis of miRNA action when their targets are located in human protein coding region. PLoS One. 2013;8(5):e63403.
Hausser J, Syed AP, Bilen B, Zavolan M. Analysis of CDS-located miRNA target sites suggests that they can effectively inhibit translation. Genome Res. 2013;23(4):604-615.
Zhang K, Zhang X, Cai Z, et al. A novel class of microRNA-recognition elements that function only within open reading frames. Nat Struct Mol Biol. 2018;25(11):1019-1027.
